Ricerca cellulare volume 31 , pagine1144–1145 ( 2021 ) Citare questo articolo
La patata con semi veri è stata per decenni il Santo Graal della coltivazione della patata; il miglioramento genetico, in particolare l’allevamento di resistenza, è rimasto indietro in questa coltura base poliploide a propagazione vegetativa. Un recente studio di Zhang et al. in Cell mostra come la genomica possa accelerare notevolmente la transizione dal tubero alla coltura da seme e aumentare i rendimenti coltivando ibridi F1 di linee consanguinee personalizzate sviluppate da risorse genetiche vegetali.
“Mamma” e “Papà” sono le prime parole che un bambino impara, ed è chiaro che madre e padre sono entità separate. La maggior parte delle specie vegetali si riproducono sessualmente come noi, ma molte di loro possono autofecondarsi in modo che la loro prole abbia un solo genitore. L’autofecondazione ripetuta porta alla consanguineità, cioè alla fissazione di tutti gli alleli in stato omozigote. La consanguineità è stata favorita dai primi agricoltori, presumibilmente perché aiutava a fissare e mantenere i tratti recessivi dell’addomesticamento, la maggior parte dei quali, come la perdita dei semi o la deiscenza dei frutti e l’adattamento a nuovi regimi di fotoperiodo, comportavano nuove mutazioni recessive. 1
Molte delle principali colture mondiali sono consanguinee, come il riso, il grano e la soia. Tuttavia, i genetisti sanno da più di un secolo che la progenie di due linee inbred supera i suoi genitori, spesso in misura piuttosto spettacolare. Le ragioni di questo vigore ibrido (noto anche come eterosi) sono rimaste sfuggenti, ma potrebbero includere la complementazione di alleli recessivi deleteri e l’azione additiva di alleli benefici dominanti. 2 I coltivatori di piante hanno combinato il meglio di entrambi i mondi: fissazione rapida e duratura degli alleli mediante consanguineità ed eterosi mediante selezione giudiziosa di gruppi eterotici di linee consanguinee da accoppiare in incroci (Fig. 1). L’aumento di sei volte della resa del mais nel XX secolo è in gran parte dovuto allo sviluppo della selezione ibrida per sostituire le varietà autoctone a impollinazione libera. 3 Lo sviluppo del riso ibrido da parte del defunto Longping Yuan lo ha reso un eroe nazionale in Cina e ha contribuito a salvaguardare la sicurezza alimentare globale. 4
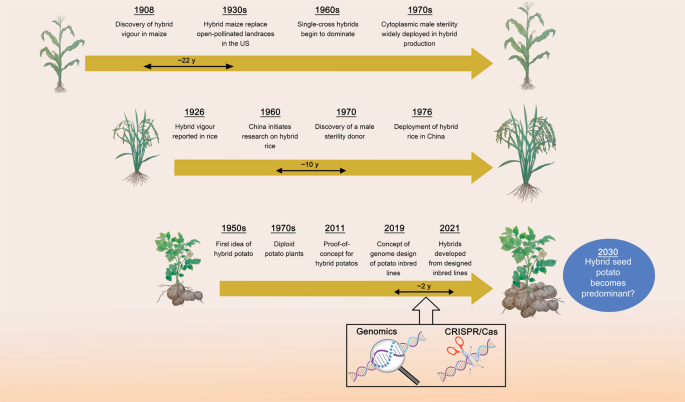
È ovvio allora chiedersi: possiamo trasformare tutte le colture in ibridi? Gli scienziati hanno individuato alcuni dei prerequisiti dell’allevamento ibrido, ad esempio lo sviluppo dei cosiddetti gruppi eterotici di linee consanguinee complementari. 5 A prima vista, la patata ( Solanum tuberosum L.) può sembrare un obiettivo improbabile per una coltura ibrida. Le varietà di patate commerciali sono autoincompatibili, il che significa che lo sviluppo di gruppi eterotici di linee consanguinee da varietà d’élite non è possibile. Inoltre, la patata è autotetraploide, poiché combina in un nucleo quattro serie di cromosomi che si ricombinano liberamente, complicando la fissazione di alleli benefici. 6 Tuttavia, queste sfide resero la prospettiva della patata con semi veri ancora più allettante. 7Poiché non esistono linee di razza pura, gli agricoltori piantano tuberi che sono cloni di alcune varietà coltivate a fini commerciali, alcune delle quali sono in uso da decenni. Dato che le popolazioni patogene si sono da tempo adattate a queste varietà, la resa delle patate può essere mantenuta solo mediante una generosa applicazione di pesticidi.
Sanwen Huang e colleghi non si sono fatti scoraggiare dalle complessità della genetica e della genomica delle patate. Hanno ideato un’intelligente combinazione di risorse genetiche vegetali, genetica tradizionale e moderno sequenziamento del genoma per dare una base razionale alla progettazione degli ibridi di patate, accelerandone allo stesso tempo il percorso. La loro pipeline in quattro fasi, dalla risorsa genetica vegetale eterozigote all’ibrido ad alto rendimento, è descritta in un recente articolo su Cell di Zhang et al. 8 e funziona come segue. Innanzitutto, Zhang et al. hanno selezionato quattro diversi cloni diploidi da banche genetiche contenenti fattori di autocompatibilità. 9 , 10I genomi di questi quattro cloni sono stati sequenziati ed è stato contato il numero di mutazioni deleterie. Zhang et al. ha optato per due cloni, denominati PG6359 ed E86-69, per inserirli nella seconda fase della loro pipeline: trovare gli selfer più promettenti. Sebbene autocompatibili e ospitassero meno varianti deleterie rispetto ad altri, i cloni selezionati erano ancora altamente eterozigoti, mascherando la maggior parte degli alleli deleteri recessivi, che si sarebbero manifestati come progenie selfed debole o sterile. Zhang et al. hanno scansionato i genomi della progenie selfed per le regioni selezionate in cui un allele parentale è molto più diffuso di quanto previsto per caso, individuando le regioni in cui l’altro genitore porta un allele deleterio. Un ostacolo notevole che hanno incontrato è stato un allele deleterio di grande effetto precedentemente sconosciuto, strettamente legato a quello desiderabileAllele Y che conferisce la polpa gialla e nutritiva del tubero. Grazie alla frequente ricombinazione sul cromosoma che ospita entrambi i geni, Zhang et al. hanno trovato una progenie che portava Y , ma era priva dell’allele deleterio. Dopo 2-4 generazioni di selfing negli consanguinei più promettenti, il successo è stato accertato mediante sequenziamento del genoma e sono state scelte linee altamente omozigoti per il quarto e ultimo passaggio. Gli ibridi F 1 sono stati sviluppati incrociando due delle loro linee inbred personalizzate e il loro vigore è stato valutato sul campo e in serra. Mini-tuberi raccolti da F 1venivano coltivati in piccoli appezzamenti. Il migliore di loro ha superato i suoi genitori di > 3 volte. Ancora più importante, la resa per ettaro prevista degli ibridi diploidi (40 t/ha) era molto vicina a quella della principale varietà di patate tetraploidi Lishu 6 (45 t/ha). E per condividere con i nostri lettori un dettaglio sincero che non è riportato nel documento: il dottor Huang ci ha assicurato in una conferenza tenuta presso il nostro istituto che i tuberi ibridi avevano un sapore davvero delizioso. Si tratta di un risultato impressionante raggiunto in un breve lasso di tempo (Fig. 1 ); tuttavia c’è ancora margine di miglioramento: l’ibrido F 1 autocompatibile ha sviluppato troppi frutti, compromettendo la resa in tuberi. E, naturalmente, l’innovazione genetica deve ancora raggiungere gli agricoltori.
Zhang et al. è tra gli articoli di genetica vegetale più stimolanti dell’ultimo decennio. Prima di tutto, è il racconto di un coltivatore che racconta come reinventare la patata come coltura da seme ibrida. Tuttavia, Zhang et al. la carta non è solo carta di patate; le sue implicazioni vanno ben oltre questa specie. La patata condivide le sue “cattive” abitudini (dal punto di vista di un allevatore) di incroci e di elevata eterozigosi con molte altre specie di colture. Possiamo utilizzare l’oleodotto delle patate per trasformare altre colture di tuberi come la manioca e l’igname o anche alberi da frutto a lunga vita in colture da seme ibride? La genomica può aiutare a trovare risorse genetiche per creare nuovi gruppi eterotici in colture ibride consolidate come mais e riso?
Riferimenti
- Dempewolf, H. et al. Cellula vegetale 24 , 2710–2717 (2012).
- Lippman, ZB e Zamir, D. Tendenze Genet. 23 , 60–66 (2007).
- Crow, JF Genetics 148 , 923–928 (1998).
- Ma, GH e Yuan, LP J. Integr. Agri. 14 , 197–205 (2015).Articolo Google Scholar
- Zhao, Y. et al. Proc. Natl Acad. Sci. Stati Uniti 112 , 15624–15629 (2015).
- Bradshaw, JE Potato Res. 60 , 171–193 (2017).
- Jansky, SH et al. Ritaglia Sci. 56 , 1412–1422 (2016).
- Zhang, C. et al. Cella 184 , 3873–3883 (2021).
- Coagulo, CR et al. Teore. Appl. Genetta. 133 , 2713–2728 (2020).
- Zhang, C. et al. Naz. Genetta. 51 , 374–378 (2019).
Finanziamento
Finanziamenti ad accesso aperto abilitati e organizzati da Projekt DEAL.
Informazioni sull’autore
Autori e affiliazioni
- Istituto Leibniz di genetica vegetale e ricerca sulle piante coltivate (IPK) Gatersleben, Seeland, GermaniaMartin Mascher, Murukarthick Jayakodi e Nils Stein
- Centro tedesco per la ricerca integrativa sulla biodiversità (iDiv) Halle-Jena-Lipsia, Lipsia, GermaniaMartin Mascher
- Centro per la ricerca sull’allevamento integrato (CiBreed), Università Georg-August di Göttingen, Göttingen, GermaniaNils Stein
Autori corrispondenti
Corrispondenza con Martin Mascher o Nils Stein .
Dichiarazioni etiche
Interessi conflittuali
Gli autori non dichiarano interessi concorrenti.
Diritti e permessi
Accesso aperto Questo articolo è concesso in licenza con una licenza internazionale Creative Commons Attribution 4.0, che consente l’uso, la condivisione, l’adattamento, la distribuzione e la riproduzione in qualsiasi mezzo o formato, a condizione che si dia il giusto credito all’autore originale e alla fonte, fornire un collegamento alla licenza Creative Commons e indicare se sono state apportate modifiche. Le immagini o altro materiale di terze parti in questo articolo sono inclusi nella licenza Creative Commons dell’articolo, se non diversamente indicato in una linea di credito al materiale. Se il materiale non è incluso nella licenza Creative Commons dell’articolo e l’uso previsto non è consentito dalle norme di legge o supera l’uso consentito, dovrai ottenere l’autorizzazione direttamente dal detentore del copyright. Per visualizzare una copia di questa licenza, visitarehttp://creativecommons.org/licenses/by/4.0/ .
